Chicos:
La pregunta es: ¿Y las fotos apa?
Mañana ya es el día. Como último día puede ser el jueves. Chequen bien la convocatoria.
Por lo pronto un poco más sobre la formación de especies y en este caso tambien la participación de la hibridación y la poliploidia.
Realiza tu resumen en el cuaderno =)
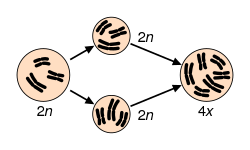
El proceso de Poliploidía por su parte, es relativamente frecuente en el reino Plantae. Si debido a procesos relacionados con la meiosis los gametos contienen más de un juego de cromosomas, al realizarse la fecundación, la célula huevo resultante será poliploide. A continuación, se requiere de una serie de condiciones: a) que la germinación ocurra, b) que la selección natural favorezca al individuo, c) que alcance la madurez sexual, d) que sea capaz de autofecundarse o que existan individuos con una mutación semejante y que puedan fecundarse; y finalmente, e) que la progenie sea viable. De este modo, tenemos que se constituye una población en la que al incrementarse el número de cromosomas, sus integrantes quedan instantáneamente aislados de los demás y pueden entonces formar una especie diferente. Se calcula que este mecanismo es responsable de la formación de nuevas especies de plantas en aproximadamente una tercera parte de las existentes en la actualidad. Ejemplos de este tipo de fenómeno lo constituyen varias de las plantas que cultiva el hombre: el trigo por ejemplo es hexaploide (6n), mientras que el tabaco es tetraploide (4n). Se ha notado que la Poliploidía con frecuencia genera plantas más vigorosas. Por ejemplo, en la fresa se ha visto que el tamaño de la planta, de la hoja y de la fruta es mayor en proporción directa al número de juegos cromosómicos presentes. Especiación alopátrica. En este proceso se pueden identificar cuatro pasos:
- Separación del acervo genético original en dos o más grupos aislados (aislamiento).
- Evolución independiente de los acervos genéticos aislados (diferenciación).
- Unión secundaria.
- Competencia entre los nuevos acervos génicos.
Aislamiento. Un grupo de poblaciones interfértiles puede separarse en dos o más grupos aislados por distintas razones. Generalmente es el cambio de ambiente físico lo que origina una reducción del territorio de las especies. La destrucción del ambiente físico por los humanos u otros tipos de disturbios pueden tener el mismo efecto. Actualmente, la separación física de poblaciones, de tal modo que estén total o parcialmente aisladas, es un fenómeno común, por lo que éste es el patrón más esperado. Las poblaciones semiaisladas o aisladas también pueden ser el resultado de una dispersión extensa, cuando un pequeño número de fundadores se establece e incrementa su densidad poblacional.
Aunque el aislamiento es necesario, no es una condición suficiente para que la especiación ocurra.
Diferenciación. Este es el proceso por el que dos poblaciones aisladas se especializan en diferentes direcciones, por lo que se hacen morfológica y/o fisiológicamente diferentes. La diferenciación no ocurre necesariamente a continuación del aislamiento, pero frecuentemente lo hace y existe amplia evidencia de que se presenta por una adaptación de la población al ambiente local.
Unión del territorio. Las fuerzas que originan una superposición de los territorios de dos poblaciones anteriormente aisladas, pueden ser similares a aquéllas que llevaron a su separación. Básicamente éstas fuerzas son cambios medioambientales que llevan a la expansión del territorio, a la migraciónde la población o a la colonización de zonas nuevas. Es posible que la evolución de las poblaciones produzca cambios fisiológicos (tal como el desarrollo de un aparato más eficiente para la dispersión de las semillas), que son directa o indirectamente responsables de la expansión. Este proceso no es una condición necesaria en el proceso de especiación. Sin embargo, cuando ocurre puede llevar a importantes interacciones entre las dos unidades anteriormente separadas.
Exclusión competitiva. Cuando dos poblaciones anteriormente separadas establecen contacto, los individuos de éstas poblaciones interactúan. Aún cuando exista un aislamiento conductual y/o genético, los individuos tendrán necesidades idénticas o similares, tales como el alimento, el espacio, sitios para vivir, etc. Si aún no hay un aislamiento reproductivo, intentarán cruzarse. La competencia puede favorecer a miembros de una población tan eficazmente, que la coexistencia puede no ser posible.
La exclusión competitiva, esto es, la eliminación de una población, es producto de la sobreposición de territorios. La población excluida puede ser la ancestral o la descendiente. Otros productos del sobrelapamiento pueden ser el desplazamiento ecológico y la especialización.
Especiación parapátrica. Este tipo de especiación puede ocurrir en especies con poca capacidad de desplazamiento, tales como plantas, animales fosorios (que tienen hábitos subterráneos, como los topos) o insectos no alados. En este proceso no existe un aislamiento geográfico muy evidente, ya que no existen barreras que les separe; el ambiente es continuo, pero su poca capacidad de movimiento impide que las poblaciones se apareen. Para que esta situación se presente se requiere de dos situaciones:
- Que sean capaces de explotar hábitats ligeramente diferentes, pero físicamente continuos.
- Que ésta capacidad de invadir nuevos hábitats les aísle reproductivamente del resto de su especie.
Este tipo de especiación probablemente no es tan común como la especiación alopátrica, pero particularmente en plantas parece ser un proceso común.
Especiación simpátrica. La mayoría de los autores hasta hace relativamente poco tiempo, consideraban a la especiación simpátrica, esto es, a la especiación sin aislamiento geográfico, como el modo más frecuente de especiación. Tal especiación se basa en dos postulados:
- El establecimiento de poblaciones nuevas de una especie en diferentes nichos ecológicos dentro del rango de dispersión de los individuos de la población parental.
- El aislamiento reproductivo de los fundadores de la población parental.
Se postula que el flujo de genes entre la población hija y la parental se inhibe más por factores intrínsecos (internos a los individuos de la población) que extrínsecos (ecológicos). En los procesos de especiación simpátrica se implica un rápido proceso de formación de especies.
Existen dos puntos básicos que deben ser aclarados: El primero es que las teorías de especiación simpátrica y geográfica (alopátrica), coinciden en la importancia que conceden a los factores ecológicos en el proceso de especiación, aunque difieran en la secuencia de los pasos. La teoría de la especiación geográfica dice que es un evento extrínseco el que separa y divide a la población original en 2 o más poblaciones y que los factores ecológicos para cada población resultante al ser distintos, juegan un papel importante en la formación de nuevas especies.
De acuerdo a la teoría de la especiación simpátrica, la fragmentación del acervo genético es originada por factores ecológicos dentro de su mismo ambiente o lugar de origen y cualquier aislamiento espacial de las poblaciones es un fenómeno posterior y secundario.
El segundo punto tiene que ver con la definición de especiación simpátrica: es el origen de mecanismos aislantes dentro del área de dispersión de las crías de un deme único (un deme es un grupo de individuos que forman una población parcialmente aislada, espacial y genéticamente de otras poblaciones similares). El tamaño de esta área está determinado, por ejemplo, en la mayoría de los organismos marinos, por la dispersión de los estados larvales. En la mayoría de los insectos se determina por la motilidad (capacidad de movimiento) del estado adulto. Un corolario necesario en cualquier teoría de especiación gradual es de que en la naturaleza deben existir algunas formas o variedades que son "especies incipientes"; es decir, que están iniciando procesos de aislamiento reproductivo. Así, se postula la existencia de razas biológicas que por lo menos en principio pueden considerarse como especies simpátricas incipientes; ejemplos de ello son las razas estacionales que se presentan en cícadas y grillos, así como las razas de hospedero tan frecuentes en parásitos como nemátodos e insectos.
Hibridación o introgresión con otras especies. Una vez que contamos con los conceptos necesarios, podemos discutir esta temática. El término híbrido puede ser definido como “el cruzamiento de individuos pertenecientes a dos poblaciones naturales distintas que tienen un contacto secundario.” Por contacto secundario queremos decir que ha existido un aislamiento geográfico previo, el cuál se rompe por la razón que sea, permitiendo encuentros entre los individuos de ambas poblaciones. Desde el punto de vista taxonómico y evolutivo (evitamos expresamente el punto de vista de floricultores y agricultores que se dedican a realizar cruzas de laboratorio), se pueden establecer 5 tipos de hibridación:
a) La cruza ocasional entre especies simpátricas, que producen híbridos inviables ya sea ecológica o conductualmente, o estériles, por lo que la cruza no trasciende.
b) La producción de híbridos mas o menos fértiles entre especies simpátricas, algunos de los cuáles sé retrocruzan con alguna o ambas especies parentales.
c) La formación de una zona secundaria de contacto donde es frecuente la hibridación, debido a que durante el aislamiento geográfico previo no se desarrollaron suficientes mecanismos de aislamiento reproductivo.
d) El rompimiento total de los mecanismos de aislamiento reproductivo entre especies simpátricas, que produce localmente formas híbridas que pueden abarcar el rango total de variabilidad de ambas especies.
e) La producción de una nueva entidad específica como resultado de la hibridación y subsiguiente duplicación del número cromosómico (aloploidìa), que únicamente se ha observado que ocurre en plantas.
Aunque la hibridación es común en plantas como pinos, encinos, castaños y muchos otros árboles, tal proceso parece ser menos frecuente en animales. No obstante, se ha documentado en animales; por ejemplo, en sapos es común que Bufo cinerea se cruce con Bufo gratiosa en zonas donde tengan contacto secundario.
Un interesante ejemplo ocurre en México entre aves del género Pipilo (ver figuras 5 y 6). El toquí de ojos rojos, Pipilo erythrophthalmus y el toquí de collar, Pipilo ocai están regularmente distribuidos como especies puras. En Oaxaca, las dos especies viven lado a lado sin mezclarse. En Puebla, aproximadamente el 16% de los individuos son híbridos, y datos similares se encuentran desde puebla hasta la costa de Nayarit, Jalisco y Michoacán. Hacia el Norte y el Este, se encuentra el toquí de ojos rojos (P. erythrophthalmus) y en el Sur y Oeste se encuentra el toquí de collar (P. ocai). La proporción de P. erythrophthalmus puros decae de Este a oeste y se incrementa la de P. ocai, igualmente decae la proporción de P. erythrophthalmus puros de norte a sur y se incrementa la de P. ocai. Es obvio que nos encontramos con un caso de hibridación tipo c.

-
-
-
-
-
-
-
-
- Figura 5. Toquí de ojos rojos (Pipilo erythrophthalmus).
-
-
-
-
-
-
-

-
-
-
-
-
-
-
-
-
- Figura 6. Distribución de Pipilo erythrophthalmus, P. ocai y las formas híbridas entre ambas especies en México.Tomado de: http://academia.cch.unam.mx/wiki/biologia3y4/index.php/Mecanismos_y_patrones_evolutivos_que_explican_la_diversidad
-
-
-
-
-
-
-
-
No hay comentarios:
Publicar un comentario